Yong Jun Kwon![]() , Jong Hee Shin
, Jong Hee Shin

1. Denning DW. Global incidence and mortality of severe fungal disease. Lancet Infect Dis 2024;24:e428-38.

2. Casalini G, Giacomelli A, Antinori S. The WHO fungal priority pathogens list: a crucial reappraisal to review the prioritisation. Lancet Microbe 2024;5:717-24.
3. Kullberg BJ and Arendrup MC. Invasive candidiasis. N Engl J Med 2015;373:1445-56.
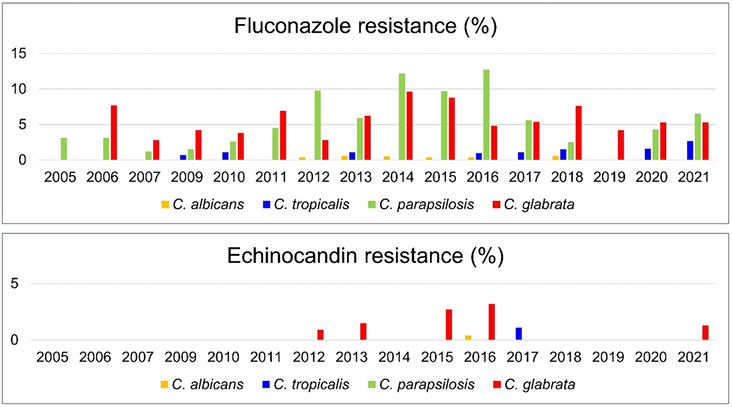
4. Won EJ, Choi MJ, Jeong SH, Kim D, Shin KS, Shin JH, et al. Nationwide surveillance of antifungal resistance of Candida bloodstream isolates in South Korean hospitals: two year report from Kor-GLASS. J Fungi 2022;8:996. 
5. Arastehfar A, Lass-Flörl C, Garcia-Rubio R, Daneshnia F, Ilkit M, Boekhout T, et al. The quiet and underappreciated rise of drug-resistant invasive fungal pathogens. J Fungi 2020;6:138.
6. Pappas PG, Lionakis MS, Arendrup MC, Ostrosky-Zeichner L, Kullberg BJ. Invasive candidiasis. Nat Rev Dis Primers 2018;4:18026.
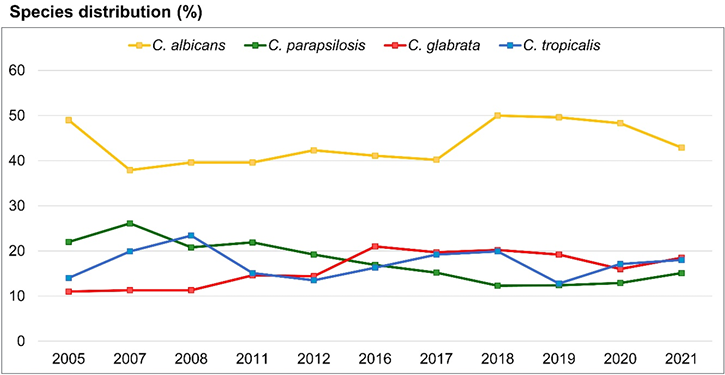
7. Guinea J. Global trends in the distribution of Candida species causing candidemia. Clin Microbiol Infect 2014;20(Suppl 6):5-10.
8. Kwon YJ, Won EJ, Jeong SH, Shin KS, Shin JH, Kim YR, et al. Dynamics and predictors of mortality due to candidemia caused by different Candida species: comparison of intensive care unit-associated candidemia (ICUAC) and Non-ICUAC. J Fungi 2021;7:597.
9. Pfaller MA, Diekema DJ, Turnidge JD, Castanheira M, Jones RN. Twenty years of the SENTRY antifungal surveillance program: results for Candida species from 1997-2016. Open Forum Infect Dis 2019;6:S79-94.
10. Won EJ, Shin JH, Choi MJ, Lee WG, Park YJ, Uh Y, et al. Antifungal susceptibilities of bloodstream isolates of Candida species from nine hospitals in Korea: application of new antifungal breakpoints and relationship to antifungal usage. PLoS One 2015;10:e0118770.
11. Jung SI, Shin JH, Song JH, Peck KR, Lee K, Kim MN, et al. Multicenter surveillance of species distribution and antifungal susceptibilities of Candida bloodstream isolates in South Korea. Med Mycol 2010;48:669-74.
12. Lee JS, Shin JH, Lee K, Kim MN, Shin BM, Uh Y, et al. Species distribution and susceptibility to azole antifungals of Candida bloodstream isolates from eight university hospitals in Korea. Yonsei Med J 2007;48:779-86.
13. Jung SI, Shin JH, Choi HJ, Ju MY, Kim SH, Lee WG, et al. Antifungal susceptibility to amphotericin B, fluconazole, voriconazole, and flucytosine in Candida bloodstream isolates from 15 tertiary hospitals in Korea. Ann Lab Med 2012;32:426-8.
14. Jang MJ, Shin JH, Lee WG, Kim MN, Lee K, Lee HS, et al. In vitro fluconazole and voriconazole susceptibilities of Candida bloodstream isolates in Korea: use of the CLSI and EUCAST epidemiological cutoff values. Ann Lab Med 2013;33:167-73.
15. Carvalhaes CG, Klauer AL, Rhomberg PR, Pfaller MA, Castanheira M. Evaluation of rezafungin provisional CLSI clinical breakpoints and epidemiological cutoff values tested against a worldwide collection of contemporaneous invasive fungal isolates (2019 to 2020). J Clin Microbiol 2022;60:e0244921.
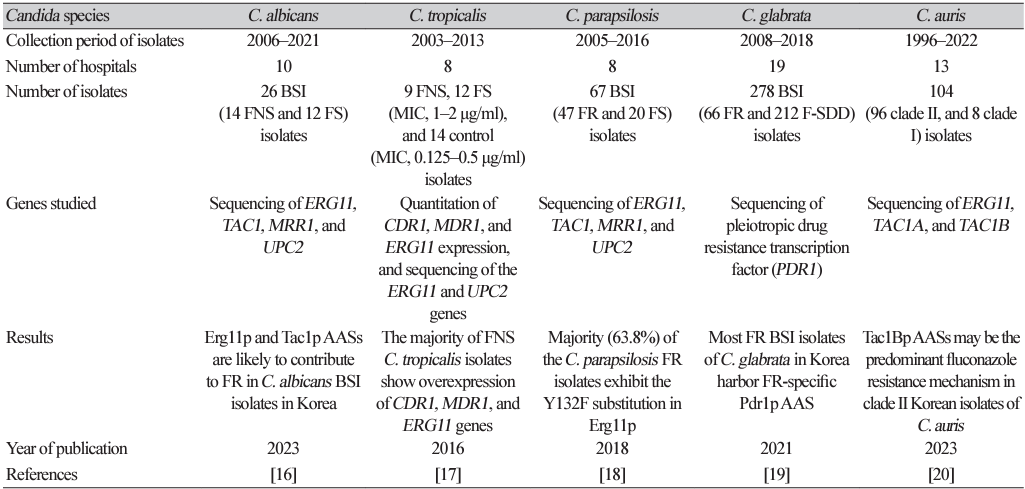
16. Choi MJ, Kwon YJ, Byun SA, Kim MN, Lee WG, Lee J, et al. Molecular and clinical features of fluconazole non-susceptible Candida albicans bloodstream isolates recovered in Korean multicenter surveillance studies. Ann Lab Med 2023;43:614-9.
17. Choi MJ, Won EJ, Shin JH, Kim SH, Lee WG, Kim MN, et al. Resistance mechanisms and clinical features of fluconazole-nonsusceptible Candida tropicalis isolates compared with f luconazole-less-susceptible isolates. Antimicrob Agents Chemother 2016;60:3653-61.
18. Choi YJ, Kim YJ, Yong D, Byun JH, Kim TS, Chang YS, et al. Fluconazole-resistant Candida parapsilosis bloodstream isolates with Y132F mutation in ERG11 gene, South Korea. Emerg Infect Dis 2018;24:1768-70.
19. Won EJ, Choi MJ, Kim MN, Yong D, Lee WG, Uh Y, et al. Fluconazole-resistant Candida glabrata bloodstream isolates, South Korea, 2008-2018. Emerg Infect Dis 2021;27:779-88.
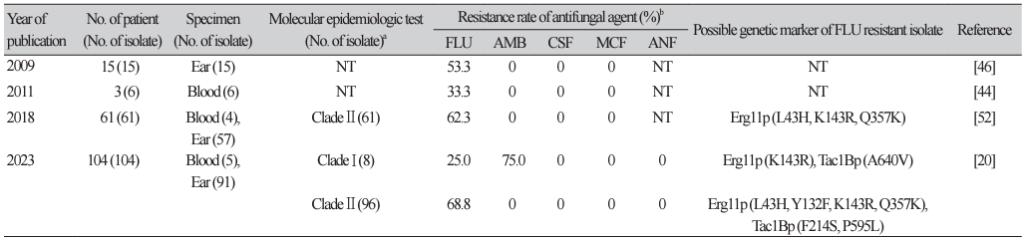
20. Byun SA, Kwon YJ, Lee GY, Choi MJ, Jeong SH, Kim D, et al. Virulence traits and azole resistance in Korean Candida auris isolates. J Fungi 2023;9:979.
21. Li J, Chen D, Yu B, He J, Zheng P, Mao X, et al. Fungi in gastrointestinal tracts of human and mice: from community to functions. Microb Ecol 2018;75:821-9.
22. Chin VK, Lee TY, Rusliza B, Chong PP. Dissecting Candida albicans infection from the perspective of C. albicans virulence and omics approaches on host-pathogen interaction: a review. Int J Mol Sci 2016;17:1643.
23. Mora Carpio AL and Climaco A. Candidemia. [Online] (last visited on 8 August 2023).
24. Swindell K, Lattif AA, Chandra J, Mukherjee PK, Ghannoum MA. Parenteral lipid emulsion induces germination of Candida albicans and increases biofilm formation on medical catheter surfaces. J Infect Dis 2009;200:473-80.
25. Shin JH, Park MR, Song JW, Shin DH, Jung SI, Cho D, et al. Microevolution of Candida albicans strains during catheter-related candidemia. J Clin Microbiol 2004;42:4025-31.
26. Escribano P, Rodríguez-Créixems M, Sánchez-Carrillo C, Muñoz P, Bouza E, Guinea J. Endemic genotypes of Candida albicans causing fungemia are frequent in the hospital. J Clin Microbiol 2013;51:2118-23.
27. Guinea J, Mezquita S, Gómez A, Padilla B, Zamora E, Sánchez-Luna M, et al. Whole genome sequencing confirms Candida albicans and Candida parapsilosis microsatellite sporadic and persistent clones causing outbreaks of candidemia in neonates. Med Mycol 2021;60:myab068.
28. Pfaller MA, Jones RN, Doern GV, Sader HS, Messer SA, Houston A, et al. Bloodstream infections due to Candida species: SENTRY antimicrobial surveillance program in North America and Latin America, 1997-1998. Antimicrob Agents Chemother 2000;44:747-51.
29. Trofa D, Gácser A, Nosanchuk JD. Candida parapsilosis, an emerging fungal pathogen. Clin Microbiol Rev 2008;21:606-25.
30. Shin JH, Kee SJ, Shin MG, Kim SH, Shin DH, Lee SK, et al. Biofilm production by isolates of Candida species recovered from nonneutropenic patients: comparison of bloodstream isolates with isolates from other sources. J Clin Microbiol 2002;40:1244-8.
31. Yamin D, Akanmu MH, Al Mutair A, Alhumaid S, Rabaan AA, Hajissa K. Global prevalence of antifungal-resistant Candida parapsilosis: a systematic review and meta-analysis. Trop Med Infect Dis 2022;7:188.
32. Arastehfar A, Hilmioğlu-Polat S, Daneshnia F, Pan W, Hafez A, Fang W, et al. Clonal candidemia outbreak by Candida parapsilosis carrying Y132F in Turkey: evolution of a persisting challenge. Front Cell Infect Microbiol 2021;11:676177.
33. Byun JH, Won EJ, Cho HW, Kim D, Lee H, Kim SH, et al. Detection and characterization of two phenotypes of Candida parapsilosis in South Korea: clinical features and microbiological findings. Microbiol Spectr 2023;11:e0006623.
34. Kim TY, Huh HJ, Lee GY, Choi MJ, Yu HJ, Cho SY, et al. Evolution of fluconazole resistance mechanisms and clonal types of Candida parapsilosis isolates from a tertiary care hospital in South Korea. Antimicrob Agents Chemother 2022;66:e0088922.
35. Arendrup MC and Patterson TF. Multidrug-resistant Candida: epidemiology, molecular mechanisms, and treatment. J Infect Dis 2017;216(suppl_3):S445-51.
36. Clinical and Laboratory Standard Institute. Performance standards for antifungal susceptibility testing of yeasts. Document M60Ed2. 2nd ed. Wayne, PA; CLSI: 2020.
37. Ferrari S, Ischer F, Calabrese D, Posteraro B, Sanguinetti M, Fadda G, et al. Gain of function mutations in CgPDR1 of Candida glabrata not only mediate antifungal resistance but also enhance virulence. PLoS Pathog 2009;5:e1000268.
38. Bassetti M, Vena A, Bouza E, Peghin M, Muñoz P, Righi E, et al. Antifungal susceptibility testing in Candida, Aspergillus and Cryptococcus infections: are the MICs useful for clinicians? Clin Microbiol Infect 2020;26:1024-33.
39. Lim HJ, Choi MJ, Byun SA, Won EJ, Park JH, Choi YJ, et al. Whole-genome sequence analysis of Candida glabrata isolates from a patient with persistent fungemia and determination of the molecular mechanisms of multidrug resistance. J Fungi 2023;9:515.
40. Goemaere B, Lagrou K, Spriet I, Hendrickx M, Becker P. Clonal spread of Candida glabrata bloodstream isolates and fluconazole resistance affected by prolonged exposure: a 12-year single-center study in Belgium. Antimicrob Agents Chemother 2018;62:e00591-18.
41. Katiyar S, Shiffrin E, Shelton C, Healey K, Vermitsky JP, Edlind T. Evaluation of polymorphic locus sequence typing for Candida glabrata epidemiology. J Clin Microbiol 2016;54:1042-50.
42. Vallabhaneni S, Cleveland AA, Farley MM, Harrison LH, Schaffner W, Beldavs ZG, et al. Epidemiology and risk factors for echinocandin nonsusceptible Candida glabrata bloodstream infections: data from a large multisite population-based candidemia surveillance program, 2008-2014. Open Forum Infect Dis 2015;2:ofv163.
43. Hwang IJ, Kwon YJ, Lim HJ, Hong KH, Lee H, Yong D, et al. Nosocomial transmission of fluconazole-resistant Candida glabrata bloodstream isolates revealed by whole-genome sequencing. Microbiol Spectr 2024;e0088324.
44. U.S. Centers for Disease Control and Prevention. 2019 Antibiotic resistance threats report. [Online] (last visited on 4 December 2024).
45. Satoh K, Makimura K, Hasumi Y, Nishiyama Y, Uchida K, Yamaguchi H. Candida auris sp. nov., a novel ascomycetous yeast isolated from the external ear canal of an inpatient in a Japanese hospital. Microbiol Immunol 2009;53:41-4.
46. Kim MN, Shin JH, Sung H, Lee K, Kim EC, Ryoo N, et al. Candida haemulonii and closely related species at 5 university hospitals in Korea: identification, antifungal susceptibility, and clinical features. Clin Infect Dis 2009;48:e57-61.
47. Lee WG, Shin JH, Uh Y, Kang MG, Kim SH, Park KH, et al. First three reported cases of nosocomial fungemia caused by Candida auris. J Clin Microbiol 2011;49:3139-42.
48. Wang Y and Xu J. Associations between genomic variants and antifungal susceptibilities in the archived global Candida auris population. J Fungi 2024;10:86.
49. Kim HY, Nguyen TA, Kidd S, Chambers J, Alastruey-Izquierdo A, Shin JH, et al. Candida auris-a systematic review to inform the world health organization fungal priority pathogens list. Med Mycol 2024;62:myae042.
50. Lockhart SR, Chowdhary A, Gold JAW. The rapid emergence of antifungal-resistant human-pathogenic fungi. Nat Rev Microbiol 2023;21:818-32.
51. Lockhart SR, Etienne KA, Vallabhaneni S, Farooqi J, Chowdhary A, Govender NP, et al. Simultaneous emergence of multidrug-resistant Candida auris on 3 continents confirmed by whole-genome sequencing and epidemiological analyses. Clin Infect Dis 2017;64:134-40.
52. Kwon YJ, Shin JH, Byun SA, Choi MJ, Won EJ, Lee D, et al. Candida auris clinical isolates from South Korea: identification, antifungal susceptibility, and genotyping. J Clin Microbiol 2019;57:e01624-18.