1. Hou K, Wu ZX, Chen XY, Wang JQ, Zhang D, Xiao C, et al. Microbiota in health and diseases. Signal Transduct Target Ther 2022;7:1-28.


2. Matijašić M, Meštrović T, Čipčić Paljetak H, Perić M, Barešić A, Verbanac D. Gut microbiota beyond bacteria-mycobiome, virome, archaeome, and eukaryotic parasites in IBD. Int J Mol Sci 2020;21:2668.
3. Kamada N, Chen GY, Inohara N, Núñez G. Control of pathogens and pathobionts by the gut microbiota. Nat Immunol 2013;14:685-90.
4. Wang H, Wei CX, Min L, Zhu LY. Good or bad: gut bacteria in human health and diseases. Biotechnol Biotechnol Equip 2018;32:1075-80.
5. LeBlanc JG, Milani C, De Giori GS, Sesma F, Van Sinderen D, Ventura M. Bacteria as vitamin suppliers to their host: a gut microbiota perspective. Curr Opin Biotechnol 2013;24:160-8.
6. Sender R, Fuchs S, Milo R. Revised estimates for the number of human and bacteria cells in the body. PLoS Biol 2016;14:e1002533.
7. Tiew PY, Mac Aogain M, Ali NAtBM, Thng KX, Goh K, Lau KJ, et al. The mycobiome in health and disease: emerging concepts, methodologies and challenges. Mycopathologia 2020;185:207-31.
8. Cui L, Morris A, Ghedin E. The human mycobiome in health and disease. Genome Med 2013;5:1-12.
9. Qin J, Li R, Raes J, Arumugam M, Burgdorf KS, Manichanh C, et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature 2010;464:59-65.
10. Kapitan M, Niemiec MJ, et al. eds. Fungi as part of the microbiota and interactions with intestinal bacteria. In: Rodrigues ML. Fungal physiology and immunopathogenesis. Switzerland; Springer, 2019:265-301.
11. Richard ML and Sokol H. The gut mycobiota: insights into analysis, environmental interactions and role in gastrointestinal diseases. Nat Rev Gastroenterol Hepatol 2019;16:331-45.
12. Yan Q, Li S, Yan Q, Huo X, Wang C, Wang X, et al. A genomic compendium of cultivated human gut fungi characterizes the gut mycobiome and its relevance to common diseases. Cell 2024;187:2969-89.
13. Bliss JM, Basavegowda KP, Watson WJ, Sheikh AU, Ryan RM. Vertical and horizontal transmission of Candida albicans in very low birth weight infants using DNA fingerprinting techniques. Pediatr Infect Dis J 2008;27:231-5.
14. Nagata R, Nagano H, Ogishima D, Nakamura Y, Hiruma M, Sugita T. Transmission of the major skin microbiota, Malassezia, from mother to neonate. Pediatr Int 2012;54:350-5.
15. Juyal D, Sharma M, Pal S, Rathaur VK, Sharma N. Emergence of non-albicans Candida species in neonatal candidemia. N Am J Med Sci 2013;5:541-5.
16. Ward TL, Dominguez-Bello MG, Heisel T, Al-Ghalith G, Knights D, Gale CA. Development of the human mycobiome over the first month of life and across body sites. MSystems 2018;3:e00140-17.
17. Zhang F, Aschenbrenner D, Yoo JY, Zuo T. The gut mycobiome in health, disease, and clinical applications in association with the gut bacterial microbiome assembly. Lancet Microbe 2022;3:e969-83.
18. Suhr MJ, Banjara N, Hallen-Adams HE. Sequence-based methods for detecting and evaluating the human gut mycobiome. Lett Appl Microbiol 2016;62:209-15.
19. Hallen-Adams H and Suhr MJ. Fungi in the healthy human gastrointestinal tract. Virulence 2017;8:352-8.
20. Otašević S, Momčilović S, Petrović M, Radulović O, Stojanović N, Arsić-Arsenijević V. The dietary modification and treatment of intestinal Candida overgrowth-a pilot study. J Mycol Med 2018;28:623-7.
21. Strati F, Di Paola M, Stefanini I, Albanese D, Rizzetto L, Lionetti P, et al. Age and gender affect the composition of fungal population of the human gastrointestinal tract. Front Microbiol 2016;7:1227.
22. Sokol H, Leducq V, Aschard H, Pham HP, Jegou S, Landman C, et al. Fungal microbiota dysbiosis in IBD. Gut 2017;66:1039-48.
23. Coker OO, Nakatsu G, Dai RZ, Wu WKK, Wong SH, Ng SC, et al. Enteric fungal microbiota dysbiosis and ecological alterations in colorectal cancer. Gut 2019;68:654-62.
24. Aminudin NI, Amran NA, Zainal Abidin ZA, Susanti D. Biotransformation of curcumin and structure-activity relationship (SAR) of its analogues: a systematic review. Biocatal Biotransformation 2023;41:1-14.
25. Bishu S, Su EW, Wilkerson ER, Reckley KA, Jones DM, McGeachy MJ, et al. Rheumatoid arthritis patients exhibit impaired Candida albicans-specific Th17 responses. Arthritis Res Ther 2014;16:1-9.
26. Zou R, Wang Y, Duan M, Guo M, Zhang Q, Zheng H. Dysbiosis of gut fungal microbiota in children with autism spectrum disorders. J Autism Dev Disord 2021;51:267-75.
27. Heisel T, Montassier E, Johnson A, Al-Ghalith G, Lin YW, Wei LN, et al. High-fat diet changes fungal microbiomes and interkingdom relationships in the murine gut. mSphere 2017;2:e00351-17.
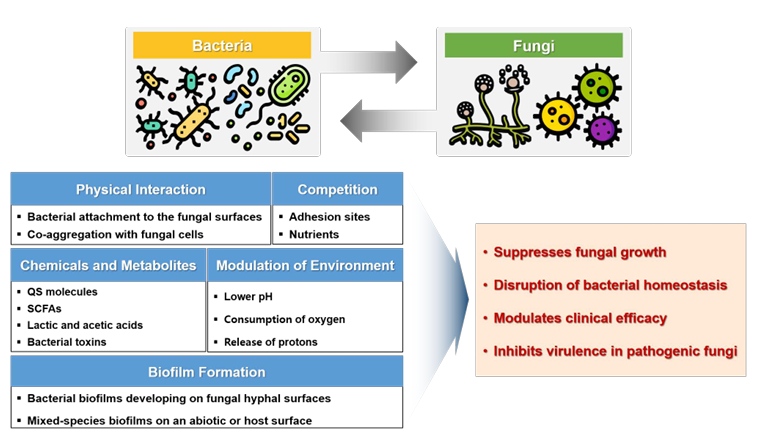
28. Lapiere A and Richard ML. Bacterial-fungal metabolic interactions within the microbiota and their potential relevance in human health and disease: a short review. Gut Microbes 2022;14:2105610.
29. MacAlpine J, Daniel-Ivad M, Liu Z, Yano J, Revie NM, Todd RT, et al. A small molecule produced by Lactobacillus species blocks Candida albicans filamentation by inhibiting a DYRK1-family kinase. Nat Commun 2021;12:6151.
30. Liu J, Huang TY, Liu G, Ye Y, Soteyome T, Seneviratne G, et al. Microbial interaction between Lactiplantibacillus plantarum and Saccharomyces cerevisiae: transcriptome level mechanism of cell-cell antagonism. Microbiol Spectr 2022;10:e01433-22.
31. Golan-Rozen N, Chefetz B, Ben-Ari J, Geva J, Hadar Y. Transformation of the recalcitrant pharmaceutical compound carbamazepine by Pleurotus ostreatus: role of cytochrome P450 monooxygenase and manganese peroxidase. Environ Sci Technol 2011;45:6800-5.
32. Martínez-Ramírez JA, Walther G, Peters FT. Studies on drug metabolism by fungi colonizing decomposing human cadavers. Part II: biotransformation of five model drugs by fungi isolated from post-mortem material. Drug Test Anal 2015;7:265-79.
33. Huang Y, Cao S, Zhang Q, Zhang H, Fan Y, Qiu F, et al. Biological and pharmacological effects of hexahydrocurcumin, a metabolite of curcumin. Arch Biochem Biophys 2018;646:317.
34. Zhang Z, Luo D, Xie J, Lin G, Zhou J, Liu W, et al. Octahydrocurcumin, a final hydrogenated metabolite of curcumin, possesses superior anti-tumor activity through induction of cellular apoptosis. Food Funct 2018;9:2005-14.
35. Zhang J, Guo H, Tian Y, Liu P, Li N, Zhou J, et al. Biotransformation of 20 (S)protopanaxatriol by Mucor spinosus and the cytotoxic structure activity relationships of the transformed products. Phytochemistry 2007;68:2523-30.
36. He X, Zhang B, Cao P, Wang H, Wu S, Wang G, et al. Biotransformation of dihydrocapsaicin by human intestinal fungi and the inhibitory effects of metabolites against LSD1. Heliyon 2022;8:e12325.
37. Zhang M, Zhang J, Wang C, Yan JK, Yi J, Ning J, et al. Biotransformation of 18β-glycyrrhetinic acid by human intestinal fungus Aspergillus niger RG13B1 and the potential anti-inflammatory mechanism of its metabolites. J Agric Food Chem 2022;70:1510415.
38. Wei X, Yao C, He X, Li J, Wang Y, Wang C, et al. Biotransformation of chenodeoxycholic acid by human intestinal fungi and the agonistic effects on FXR. Phytochemistry 2024;224:114162.
39. Bowman MJ, Jordan DB, Vermillion KE, Braker JD, Moon J, Liu ZL. Stereochemistry of furfural reduction by a Saccharomyces cerevisiae aldehyde reductase that contributes to in situ furfural detoxification. Appl Environ Microbiol 2010;76:4926-32.
40. Schoch CL, Seifert KA, Huhndorf S, Robert V, Spouge JL, Levesque CA, et al. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. PNAS 2012;109:6241-6.
41. Nash AK, Auchtung TA, Wong MC, Smith DP, Gesell JR, Ross MC, et al. The gut mycobiome of the Human Microbiome Project healthy cohort. Microbiome 2017;5:1-13.
42. Xie Z and Manichanh C. FunOMIC: pipeline with built-in fungal taxonomic and functional databases for human mycobiome profiling. Comput Struct Biotechnol J 2022;20:3685-94.
43. Langsiri N, Worasilchai N, Irinyi L, Jenjaroenpun P, Wongsurawat T, Luangsa-Ard JJ, et al. Targeted sequencing analysis pipeline for species identification of human pathogenic fungi using long-read nanopore sequencing. IMA fungus 2023;14:18.
44. Lewis JD, Chen EZ, Baldassano RN, Otley AR, Griffiths AM, Lee D, et al. Inflammation, antibiotics, and diet as environmental stressors of the gut microbiome in pediatric Crohn’s disease. Cell Host Microbe 2015;18:489-500.
45. Lee EH, Kim H, Koh JH, Cha KH, Lee KK, Kim WU, et al. Dysbiotic but nonpathogenic shift in the fecal mycobiota of patients with rheumatoid arthritis. Gut Microbes 2022;14:2149020.
46. Wang H, Wu H, Li KD, Wang YY, Huang RG, Du YJ, et al. Intestinal fungi and systemic autoimmune diseases. Autoimmun Rev 2023;22:103234.
47. Lin Y, Lau HCH, Liu Y, Kang X, Wang Y, Ting NLN, et al. Altered mycobiota signatures and enriched pathogenic Aspergillus rambellii are associated with colorectal cancer based on multicohort fecal metagenomic analyses. Gastroenterology 2022;163:908-21.
48. Wang T, Fan C, Yao A, Xu X, Zheng G, You Y, et al. The adaptor protein CARD9 protects against colon cancer by restricting mycobiota-mediated expansion of myeloid-derived suppressor cells. Immunity 2018;49:504-14.e4.
49. Chen Y, Chen Z, Guo R, Chen N, Lu H, Huang S, et al. Correlation between gastrointestinal fungi and varying degrees of chronic hepatitis B virus infection. Diagn Microbiol Infect Dis 2011;70:492-8.
50. Mukherjee PK, Chandra J, Retuerto M, Tatsuoka C, Ghannoum MA, McComsey GA. Dysbiosis in the oral bacterial and fungal microbiome of HIV-infected subjects is associated with clinical and immunologic variables of HIV infection. PLoS One 2018;13:e0200285.
51. Jha AK, Uppal B, Chadha S, Bhalla P, Ghosh R, Aggarwal P, et al. Clinical and microbiological profile of HIV/AIDS cases with diarrhea in North India. J Pathog 2012;2012:971958.
52. Mar Rodríguez M, Pérez D, Javier Chaves F, Esteve E, Marin-Garcia P, Xifra G, et al. Obesity changes the human gut mycobiome. Sci Rep 2015;5:14600.
53. Shah S, Locca A, Dorsett Y, Cantoni C, Ghezzi L, Lin Q, et al. Alterations of the gut mycobiome in patients with MS. EBioMedicine 2021;71:103557.
54. Zhang X, Pan L, Zhang Z, Zhou Y, Jiang H, Ruan B. Analysis of gut mycobiota in first-episode, drug-naïve Chinese patients with schizophrenia: a pilot study. Behav Brain Res 2020;379:112374.
55. Severance EG, Alaedini A, Yang S, Halling M, Gressitt KL, Stallings CR, et al. Gastrointestinal inflammation and associated immune activation in schizophrenia. Schizophr Res 2012;138:48-53.